We certify that we have
read this thesis and that in our opinion,
it is satisfactory in
scope and quality as a thesis for the degree of Master of Science in
Microbiology.
|
|

PLASMID
CONTROL OF SYMBIOTIC PROPERTIES
IN
Rhizobium fredii
A
THESIS SUBMITTED TO THE GRADUATE DIVISION OF THE
UNIVERSITY
OF HAWAII IN PARTIAL FULFILLMENT
OF
THE REQUIREMENTS FOR THE DEGREE OF
MASTER
OF SCIENCE
IN
MICROBIOLOGY
AUGUST,
1986
By
Maria
Luz Cases
Thesis
Committee
B.
Ben Bohlool, Chairman
Leslie
R. Berger
Francoise
M. Robert
We certify that we have
read this thesis and that in our opinion,
it is satisfactory in
scope and quality as a thesis for the degree of Master of Science in
Microbiology.
|
|
ACKNOWLEDGEMENTS
I would like to express my gratitude to Dr.
Ben Bohlool for the opportunity to do research under his tutelage and financial
support. I would also like to thank the
members of my committee for their constructive suggestions in the preparation
of this thesis.
I am indebted to Mark Kingsley for his
valuable contributions in all aspects of this research by his technical
guidance, helpful comments find friendship.
I would also like to acknowledge Dr. Duane Bartholomew of the Department
of Agronomy for the use of the growth chambers.
Lastly, I would like to say a very special
thank you to my parents and family for their support and encouragement, and to
my best friend, Thomas George, for his help, patience, and understanding.
The research was funded in part by grants
58-9AHZ-2-670 from
the U. S. Department of Agriculture and
DAN-5542-G-SS-2095-00 from the U. S. Agency for International Development.
ABSTRACT
Large indigenous plasmids are a common
feature in members of the genus Rhizobium, and their involvement in the
control of symbiotic functions has been established. Studies on these rhizobial plasmids, called pSym plasmids, make
up much of what is understood about the molecular and genetic basis of the N2-fixing
symbiosis. The soybeans, generally
nodulated by slow-growing rhizobia, are an economically important crop but
genetic studies about this group of rhizobia have made little progress because
of their slow growth rate and the absence of identifiable plasmid-associated
symbiotic functions. An approach to
understanding the symbiosis in the soybean system is to use as a genetic model,
R. fredii, the fast-growing rhizobia that nodulate soybeans.
In this study, five strains of R. fredii
were examined for the presence of indigenous plasmids. To determine if symbiotic functions are
controlled by genes on these plasmids, the strains were subjected to
plasmid-curing treatments. The effect
of a pSym plasmid from a heterologous species on R. fredii gene
function was studied by the introduction of the R. leguminosarum
pSym plasmid, pJB5JI.
The results of this study show that high
molecular weight plasmids that are involved in determining symbiotic functions,
as well as cryptic plasmids, are an integral part of the genetic make-up of R.
fredii. These strains can
receive and maintain p7B5JI, a pSym plasmid from another Rhizobium
species. However, the pJB5JI plasmid
genes are not expressed in the R. fredii genetic background. The introduction of the plasmid did not
enable any of the R. fredii transconjugants to nodulate peas, nor
did it restore the ability of the plasmid-cured transconjugants to nodulate
soybeans. The presence of the plasmid
affected the expression of the R. fredii symbiotic genes,
resulting in different levels of symbiotic effectiveness.
TABLE
OF CONTENTS
ACKNOWLEDGEMENTS
.................................... 3
ABSTRACT
............................................
4
LIST
OF TABLES ...................................... 7
LIST
OF FIGURES ..................................... 8
CHAPTER 1. INTRODUCTION ............................ 9
CHAPTER
2. MATERIALS AND METHODS
................... 22
Bacterial
Strains ....................... 22
Maintenance
of Cultures ................. 22
Assessment
of Culture Purity and
identification
of strains ............... 22
Plasmid
Profile Analysis ................ 25
Development
of Mutants .................. 26
Bacterial
Matings ....................... 28
Plant
Infection Tests ................... 29
CHAPTER
3. RESULTS
................................. 32
Plasmid
Profiles ........................ 32
Isolation
of Plasmid-Cured Mutants ...... 38
Transfer,
Expression and Maintenance
of
pSym Plasmid ......................... 41
CHAPTER
4. DISCUSSION
.............................. 58
LITERATURE
CITED.....................................
66
LIST
OF TABLES
Table Page
1 Characteristics and sources of
Rhizobium
strains used in the study ...........
23
2 Acid production and FA reactions of
R. fredii
and R. leguminosarum strains ........ 33
3 Intrinsic antibiotic resistance
patterns
of R. fredii
and R. leguminosarum strains ..... 34
4 Phage-typing patterns of R. fredii
and
R. leguminosarum
strains ...................... 35
5 Symbiotic properties of parent and
plasmid-
cured R.
fredii strains on soybeans ...........
42
6 Frequency of natural kanamycin
resistance
in R. fredii
strains .......................... 44
7 Frequency of transfer of kanamycin
resistance
marker in
crosses between R. leguminosarum 6015
(pJB5JI) and R.
fredii ........................
45
8 Presence or absence of the plasmid
pJB5JI
and symbiotic
properties of R. fredii
transconjugants................................ 46
9 Frequency of transfer of kanamycin
resistance
marker in
backcrosses between R. fredii
transconjugants
and R. leguminosarum 6015 .....
50
10 Summary of the pJB5JI donor, plasmid
content, and
symbiotic properties of
R. leguminosarum
6015 transconjugants ......... 54
11 Nodulation and nitrogen fixation by
isolates of R.
fredii transconjugants after
first passage
on Peking soybeans .............. 55
12 Nodulation and nitrogen fixation by
isolates of R.
fredii transconjugants after
second passage
on Peking soybeans ............. 57
LIST
OF FIGURES
Figure Page
1 Agarose gel electrophoresis of plasmid
DNA from R.
fredii ............................
37
2 Plasmid profiles of R. fredii
cured
mutants and
parent strains .................... 40
3 Plasmid profiles of USDA 205-A05
transconjugants
and reference strains ......... 49
4 Plasmid profiles of R. leguminosarum
6015 and its
transconjugants .................. 52
CHAPTER
1
INTRODUCTION
The
rhizobia are Gram-negative soil bacteria belonging to the genus Rhizobium
which fix nitrogen in a symbiotic association with legumes. Establishment of the N2-fixing
symbiosis starts with the colonization, recognition and invasion of the plant root by free-living rhizobia in the
soil, followed by a series of steps that result in the formation of a
nodule. It is in these nodules that the
rhizobia differentiate into bacteroids and reduce atmospheric nitrogen to
ammonia with the enzyme nitrogenase.
In
the symbiosis, both the plant host and the bacterial endosym-biont play
particular roles that are regulated by gene expression. The frequency with which certain symbiotic
properties such as infectiveness and effectiveness were lost, and the stimulation
of this loss by treatments known to affect plasmids has led to the suggestion
that symbiotic properties are plasmid-borne.
Indigenous plasmids in rhizobia
The
presence of plasmids with molecular weights higher than 100 megadaltons (Mdal)
is a general feature in the family Rhizobiaceae. This has been correlated with some biological effects of the
bacterial symbiont on the plant, including the induction of crown gall by Agrobacterium
and the establishment of N2-fixing symbioses by Rhizobium and
Bradyrhizobium. In rhizobia, the
plasmids have also been found to bear genes for the production of the melanin
pigment (Beynon, et al., 1980), bacteriocins (Hirsch, 1979), cell wall
polysaccharides (Prakash, 1980) and uptake hydrogenase (Brewin et al.,
1980c). However, the function of most
of the DNA in these plasmids remains unknown.
Strains
of both the fast-growing Rhizobium and the slow-growing Bradyrhizobium
typically harbor one to several plasmids.
A great percentage of their DNA is in the form of large, low copy number
(1-3 per cell) plasmids, representing at least 5 to 20 per cent of the total
genomic information (Beringer et al., 1980).
In
terms of physical evidence, many indigenous plasmids were isolated by alkaline
denaturation-phenol extraction and visualized by gel electrophoresis (Casse et
al., 1979; Hirsch et al., 1980; Nuti et al., 1977; Prakash et al., 1980). The larger megaplasmids were detected by the
more sensitive Eckhardt direct-lysis method of gel electrophoresis (Denarie et
al., 1981; Eckhardt, 1978; Rosenberg et al., 1982;).
Large
plasmids were found in Rhizobium leguminosarum, Rhizobium trifolii,
Rhizobium phaseoli, and Rhizobium meliloti (Beynon
et al., 1980b; Casse et al., 1979; Denarie et al., 1981; Hirsch et al., 1980;
Nuti et al., 1977; Prakash, 1980). In R.
meliloti, different isolation procedures revealed two classes of
plasmids. One class consisted of medium
range 90- to 200-Mdal plasmids (Casse et al., 1979) and a second class
consisted of plasmids with sizes greater than 300 Mdal, referred to as
megaplasmids (Banfalvi et al., 1981; Denarie et al., 1981; Rosenberg et al.,
1981). By means of electron microscopy, Burkhardt and Burkhardt (1984)
estimated their size to be 1000 Mdal.
In
the case of slow-growing rhizobia, Gross et al. (1979) examined Bradyrhizobium
japonicum isolated from alkaline soils and observed large plasmids. Other strains from various geographical
origins ware examined by Masterson et al. (1982) and all of those examined
contained at least one large plasmid, with sizes ranging from 118 to 1915
Mdal. They concluded that the presence
of plasmids is a consistent feature of most B. japonicum strains.
Evidence
has been presented that in the endosymbiotic bacteroids, the large plasmids are
extensively transcribed. Krol et al.
(1980) studied R. leguminosarum and found that in contrast to the
bacteroids, there was no detectable transcription of the plasmid in broth
cultures, showing that under these conditions, plasmid genes are not
expressed. They suggested that the
large rhizobium plasmids contain not only the genes that control N2-fixation
but also the genes which are functional during the differentiation of bacteria
into bacteroids.
Plasmid involvement in symbiotic nitrogen
fixation
Physical
and genetic studies have now established that, at least in fast-growing Rhizobium
species, the majority of the genes that control symbiotic functions such as
host-range specificity, nodulation, and N2-fixation are usually on a
plasmid referred to as the pSym plasmid.
In
several fast-growing Rhizobium species, genes controlling symbiotic
functions have been localized on large plasmids ranging in size from 130 Mdal
in R. leguminosarum (Hirsch et al., 1980) to more than 450 Mda1
in R. meliloti (Denarie et al., 1981). In slow-growing rhizobia such as B. japonicum,
plasmids controlling symbiotic functions have not been identified and the
symbiotic genes are presumed to be located on the chromosome.
Host
range specificity
Host
selectivity at the early stages of infection is a striking feature of the
rhizobia-plant interaction. With one
known exception (Parasponia), the rhizobial plant hosts are members of
the family Leguminosae, and within this family, groups of plants are nodulated
by particular Rhizobium species.
There is evidence that the pSym plasmid carries genes determining
host-range specificity (hsn).
Brewin
et al. (1981) reviewed the role of Rhizobium plasmids in host
specificity, and concluded that, at least for the three closely related
species, R. leguminosarum, R. trifolii, and R. phaseoli, host-range is a
plasmid-determined trait.
Plasmid-controlled
host specificity in R. leguminosarum was first described by
Johnston et al. (1978). In the study, a
R. leguminosarum plasmid, pJB5JI, was transferred to R. trifolii
and R. phaseoli. All
transconjugants were capable of forming nodules on peas in addition to their
normal hosts. By cloning the nodulation
genes of the plasmid, Downie et al. (1983) showed that host specificity is
determined by a DNA sequence of no more than 10 kilobases (kb).
In
R. trifolii, mutagenesis of host-specific nodulation genes in the
pSym plasmid by the transposon Tn5, a short segment of DNA coding for the
antibiotic kanamycin and which is capable of inserting within a genome,
resulted in mutants with altered host-range ability. The mutants showed either poor nodulation on clover or none at
all (Djordjevic et al., 1985). A 14-kb
fragment of the pSym plasmid was mobilized and conferred clover-specific
nodulation on another strain of R. trifolii that was cured of the
pSym plasmid, as well as on Agrobacterium tumefaciens, a member
of another genus in Rhizobiaceae (Schofield et al., 1984).
Particular
host-range characteristics within a cross inoculation group were also found to
be plasmid-controlled. The strain R.
leguminosarum TOM, which nodulates the primitive pea cultivar
Afghanistan, was able to transfer this trait at low frequencies to a strain
that does not normally nodulate this cultivar.
The transfer of cultivar specificity was associated with the transfer of
a 160-Mdal plasmid designated pRL5JI (Brewin et al., 1980).
Other
reports gave evidence for the presence of host-range determinants on pSym
plasmids (Appelbaum et al., 1985; Beynon et al., 1980; Djordjevic et al., 1983;
Hooykaas et al., 1981; Kondorosi et
al., 1982; Morrison et al., 1983).
Nodulation
Successful
interaction of a rhizobia with their plant host results in the development of
an organized structure, the root nodule.
The early stages of nodule formation include root hair colonization and
adhesion, root hair curling, infection thread development, cortical cell
multiplication, release of rhizobia in the host cells, and proliferation of
bacteria within the plant cells. The later
stages encompass events such as bacteroid differentiation, host cell
enlargement, and the start of nitrogen fixation (Verma and Long, 1983).
The
role of plasmids in the control of nodule formation has been demonstrated by
studies on plasmid-cured mutants, transposon mutagenesis, and plasmid transfer
experiments.
The
association of plasmid loss and a non-nodulatiog (Nod-) phenotype
provided early evidence of plasmid involvement in nodulation. Higashi (1967) found that the ability of R.
trifolii to nodulate clover was lost at high frequency following
treatment with acridine orange, a chemical agent known to cause the elimination
of plasmids (Clowes et al., 1965; Parijkaya, 1973).
Zurkowski
and Lorkiewicz (1976, 1978) correlated plasmids in R. trifolii
with nodulation, by showing that Nod- mutants resulting from a
prolonged treatment at high temperature were due to either loss of plasmid DNA
or internal deletions in the plasmid.
Zurkowski (1982) reported that at high temperature, DNA synthesis stops
while protein synthesis continues, leading to the formation of enlarged cells,
and the loss of the plasmid during cell division. The transfer of the plasmid into the Nod- strains
which had lost the ability to attach to the root hair surface, converted them
to a Nod+ phenotype (Zurkowski, 1981). The absence of the symbiotic function that was associated with
plasmid loss and its restoration with plasmid transfer, point to the
involvement of the plasmid in the nodulation process.
Similarly,
prolonged heat treatment of R. leguminosarum resulted in Nod-
strains that exhibited modified surface properties (Prakash et al., 1980). The mutants lost the ability to agglutinate
in the presence of pea lectin, a plant protein that recognizes and binds with
specific carbohydrate components of the bacterial cell surface. Lectin binding is a function that has been
correlated with host specificity (Bohlool and Schmidt, 1974; Dazzo and Hubbel,
1975).
In
R. meliloti, the early functions in the infection process are
reported to be on a megaplasmid (Rosenberg et al., 1981). Previously, Palomares et al. (1978) had
shown that in R. meliloti, extrachromosomal DNA was responsible
for polygalacturonase, a key enzyme originally thought to be involved in the
early infection process (Ljunggren and Fahraeus, 1961). Long et al. (1982) cloned the nod genes
which complement the nodulation defect of a Nod- R. meliloti
mutant and assigned the nodulation function to a region of the 8.7-kb EcoRI
fragment on the megaplasmid.
Although
the nodulation process is characterized by a high degree of specificity, some
of the genes involved in the early steps of nodulation were found to be
conserved and common across different rhizobia. A proposed genetic model is that there is a core of nodulation-specific
genes that are essentially the same in different Rhizobium species and
that host range is determined by ancillary host range genes (Downie et al.,
1983).
Kondorosi
et al. (1984) identified DNA regions of a R. meliloti megaplasmid
carrying nod genes involved in root hair curling, an early step in nodule
formation. The genes, referred to as
the common nod genes, are active in a wide range of plant hosts and
mutations in these can be complemented by pSym plasmids from other Rhizobium
species such as R. leguminosarum. The genes nodABCD, which are clustered in the pSym plasmid
have been identified in several species of Rhizobium (Banfalvi et al.,
1981; Djordjevic et al., 1985; Fischer et al., 1985).
The
functional conservation of nod genes in the fast-growing strains may also
extend to the slow growers. When DNA
fragments from Bradyrhizobium sp. (Parasponia) were introduced
into a Nod- R. meliloti strain, nodulation ability was
restored (Marvel et al., 1984). Noti et
al. (1985), using a DNA region from Bradyrhizobium sp. (Vigna),
reported similar findings.
Nitrogen
fixation
Johnston
et al. (1978) demonstrated that the transfer of a R. leguminosarum
plasmid into a Fix- strain restored its normal symbiotic function,
implying that the genes that control the ability to fix nitrogen are located on
plasmids. Furthermore, the transfer of
the R. leguminosarum bacteriocinogenic plasmid pRLlJI into
symbiotic mutants, including nodulation-deficient ones restored them to Fix+
phenotypes (Brewin et al., 1980a). The
location of the genes involved in N2-fixation was confirmed by
hybridization with a recombinant DNA clone carrying the nitrogenase (nif) genes
from Klebsiella pneumoniae.
The K. pneumoniae nifD and nifH genes, which encode two of
the three nitrogenase subunit polypeptides, hybridizes to DNA restriction
fragments of many diverse N2-fixing bacteria, indicating the
conservation of the nif genes at the DNA sequence level (Ruvkun and Ausubel,
1980a). Hybridization experiments have
established the presence of nif genes in R. leguminosarum (Nuti et
al., 1979), R. phaseoli (Hombrecher et al., 1981), R. trifolii
(Hooykaas et al., 1981; Schofield et al., 1985), R. meliloti
(Banfalvi et al., 1981; Rosenberg et al., 1981), and a fast-growing cowpea Rhizobium
(Morrison et al., 1983). Homology
studies between the pSym plasmids of diverse fast-growing Rhizobium
species showed that a specific DNA sequence which carries the structural genes
for nitrogenase is highly conserved in R. leguminosarum, R.
trifolii, and R. phaseoli (Prakash et al., 1981).
Slow-growing
rhizobia, including B. japonicum have not been shown to carry nif
sequences on plasmid DNA (Masterson et al., 1982).
Linkage
of genes
The
genes for nodulation and N2-fixation as well as for other functions
are linked on a plasmid in most Rhizobium strains. In many cases, the pSym plasmid found to
bear nif sequences was also known to encode for nodulation and host-range
specificity, and for other functions.
Tn5
insertions in a R. leguminosarum plasmid (Buchanan-Wollaston et
al., 1980) and in a R. meliloti megaplasmid (Meade et al., 1982)
could produce both Nod- and Fix- phenotypes. In studies of the R. meliloti
megaplasmid, it was shown that in a large number of spontaneous Nod-
mutants, a deletion of the megaplasmid occurred with the concommitant loss of
the sequence homologous to nif.
By analyzing such deletions, Banfalvi et al. (1981) and Rosenberg et al.
(1981) deduced close linkage of the nod and nif loci. Physical and genetic data have confirmed
that the nod genes are located within 30 kb of the nif genes on
the megaplasmid (Long et al., 1982).
Hybridization
experiments showed that nodulation and nitrogen fixation genes are found in the
same plasmid in R. leguminosarum (Hombrecher et al., 1981), R.
phaseoli (Lamb et al., 1982), R. trifolii (Hooykaas et
al., 1981) and a fast-growing cowpea Rhizobium (Morrison et al.,
1983). The molecular linkage map of the
nitrogenase and nodulation genes was constructed in R. trifolii
by Schofield et al. (1983). Hooykaas et
al. (1981) reported that the R. trifolii pSym plasmid not only
determines host specificity for clover, but also controls other steps in
nodulation and N2-fixation.
In
crosses between R. leguminosarum and R. trifolii,
host-range specificity is cotransferred with other nodulation loci (Djordjevic
et al., 1983; Downie et al., 1983; Hombrecher et al., 1984; Schofield et al.,
1984).
Close
linkage with the symbiotic genes on the pSym plasmids is shown by genes that
control other functions. Beynon et al.,
(1980b) observed that spontaneous plasmid deletions eliminated both melanin
producticn and the ability to nodulate Phaseolus beans and suggested
that the genes involved in the control of both functions are on a single
plasmid. Brewin et al. (1980c)
identified a R. leguminosarum plasmid which not only carried nif
and nod genes but also specified the genes controlling the production of uptake
hydrogenase, an enzyme that catalyzes the oxidation of the hydrogen liberated
during nitrogen reduction.
Expression of the pSym plasmid in different
species
The
transfer of indigenous Rhizobium plasmids has been reported earlier by
Cole and Elkan (1973), Higashi (1967), Hirsch (1979), and Johnston et al.
(1978). The pSym plasmids in some
strains of R. trifolii, R. leguminosarum, and R.
phaseoli are self-transmissible, but the pSym megaplasmids of R. meliloti
are not. They have been transferred to
other strains by recombination with a transmissible plasmid (Brewin et al.,
1980a), by cointegration with RP4 (Kowalczuk et al., 1981; Scott and Ronson,
1982), and by introduction of a mobilization site (Simon, 1983).
The
transfer of plasmids coding for symbiotic functions to different species often
results in variable expression.
Hooykaas et al. (1981) observed that the pSym plasmids of R. leguminosarum
and R. trifolii expressed symbiotic properties fully when
transferred between these two strains.
However, when transferred to R. meliloti, the
transconjugants formed nodules on the host plants but did not fix
nitrogen. The same results were shown
when these pSym plasmids were transferred to Agrobacterium tumefaciens. An examination of the nodules by electron
microscopy showed the presence of numerous infection threads and a
peribacteroid membrane but not the presence of bacteroids. When the nif and nod genes
from the pSym megaplasmid of R. meliloti were mobilized by RP4
and transferred to A. tumefaciens, the transconjugants induced
root deformations which cytologically resembled real nodules in alfalfa
(Truchet, 1984).
The
transfer of the R. leguminosarum plasmid pJB5JI to a Nod-
R.
trifolii strain resulted in transconjugants that formed effective
nodules on peas but no nodules on clover.
However, the transconjugant reisolates from the pea nodules were able to
nodulate clover (Sadowsky,, 1983; Sadowsky and Bohlool, 1985).
Djordjevic
et al. (1983) found that the transfer of the plasmid pBRIAN which encodes
clover specificity, to plasmid-cured R. meliloti strains did not
result in the ability of R. meliloti to nodulate clover.
According
to Djordjevic et al. (1982) and Christensen and Schubert (1983),
incompatibility and instability of the pSym plasmids may be factors that
contribute to the variable expression of symbiotic genes in different hosts.
Objectives of present study
Considerable
progress has been made in understanding the rhizobial genes involved in
symbiotic N2-fixation with the fast-growing strains. Genetic studies
of the symbiotic relationship between legumes and rhizobia have relied largely
on R. leguminosarum, R. trifolii, R. phaseoli,
and R. meliloti. This is
mainly due to their fast growth rate, ease of handling and lysis as well as
their suitability for recombinant DNA techniques.
Progress
has been hindered in the genetic analysis of the slow-growing strains due to
the absence of identifiable plasmid-associated symbiotic genes. Although the organization of structural
genes for nitrogenase is understood in the slow-growing B. japonicum,
little is known about the other genes involved in symbiotic nitrogen
fixation. At present, there are no
reliable methods of gene transfer systems in the slow-growing species.
Kuykendall (1979) and Pilacinski and Schmidt (1981) have been successful in
introducing Pl group plasmids (R68.45 and RP4 originally from Pseudomonas
and E. coli) into B. japonicum and other cowpea
rhizobia but with a very low frequency.
The genetic analysis of the slow-growing rhizobia is further complicated
by the observed instability of their genome, as evidenced by the spontaneous
indigenous rearrangements between the chromosome and plasmid DNA (Berry and
Atherly, 1984).
The
dearth of knowledge in this area is unfortunate because these rhizobia nodulate
agronomically important legumes. They
are also bio-chemically well characterized and can routinely be shown to reduce
significant amounts of acetylene as free-living bacteria (Keister, 1975).
The
fast-growing strains of soybean-nodulating rhizobia isolated from the People's
Republic of China (Keyser et al.,(1982) present an alternative model for
studying the genetic control of the N2-fixing symbiosis in the
soybean system. These strains represent
a class of rhizobia intermediate between classical fast- and slow-growing
types. They resemble the fast-growing
species physiologically and nodulate Glycine soja, the ancestral
soybean, and Glycine max.
However, they generally form ineffective nodules on the commercial
soybean cultivars.
Members
of the group, classified as Rhizobium fredii (Scholla and Elkan,
1984), harbor high-molecular weight plasmids.
The strains examined contain 1-3 plasmids with sizes up to 200 Mdal
(Masterson et al., 1982; Sadowsky and Bohlool, 1985) and most were also found
to contain plasmids that are larger than 450 Mdal (Appelbaum et al., 1984;
Broughton et al., 1984; Heron and Pueppke, 1984).
Both
nodulation and nitrogenase genes have been shown to be carried on the 150-200
Mdal plasmids, with some exceptions which may carry the symbiotic genes on the
chromosome (Broughton et al., 1984; Masterson et al., 1982). A single plasmid of about 200 Mdal has
homology to a cloned DNA fragment containing the nifD and nifH
genes of R. meliloti in most of the strains examined (Masterson
et al., 1982). In two strains, plasmid
DNA hybridizes to both R. meliloti nif and nod
sequences (Prakash and Atherly, 1984).
Loss of a 200-Mdal plasmid from one strain results in loss of the
ability to nodulate soybeans (Sadowky and Bohlool, 1983). Mobilization of a 200-Mdal plasmid from USDA
191 to other Rhizobium strains that neither nodulate soybeans nor form
effective nodules on certain cultivars enabled the transconjugants to form
nodules or nodule-like structures on soybeans (Appelbaum et al., 1985). The symbiotic properties of the
transconjugants indicate that both soybean specificity for nodulation and
cultivar specificity for nitrogen fixation are plasmid-encoded in R. fredii
(Appelbaum et al., 1985).
Sadowsky
and Bohlool (1985) used these soybean rhizobia and R. trifolii
strains as recipients of the R. leguminosarum plasmid pJB5JI, and
reported the differential expression of the plasmid in the different genetic
backgrounds.
Masterson
et al. (1985) demonstrated the conservation of DNA sequences of the symbiotic
genes within and between B. japonicum and R. fredii
strains. The structural
organization of the symbiotic genes appears highly conserved in the R. fredii
and B. japonicum strains examined regardless of the plasmid or
chromosomal location of the genes.
A
second group of R. fredii strains that was isolated (Dawdle,
1985) is especially attractive for genetic studies because the strains form
effective nodules on the commercial soybean cultivar Williams as well as
Peking. This group is similar to USDA
191, the only strain from the first set of R. fredii (Keyser et
al., 1982) known to form Fix+ nodules on commercial cultivars
(Hattori and Johnson, 1984). It was
also shown that these strains were highly competitive on two soybean cultivars
(Williams and Ai Jiao Zao) in the soil where they naturally occurred, and under
certain conditions, are effective as inoculum strains (Dawdle, 1985).
By
using the soybean system as a model, the purpose of this research is to
understand the genetic control of the Rhizobium-legume symbiosis. With this broad aim, the following
objectives were formulated:
1. To examine the fast-growing R. fredii
strains for the presence of large indigenous plasmids;
2. To determine whether symbiotic genes are
carried on these plasmids; and,
3.
To study the transfer, expression and maintenance of pSYM plasmids from another
species.
CHAPTER
2
MATERIALS
AND METHODS
Bacterial strains
The
Rhizobium strains used in this study, their relevant characteristics,
and sources are listed in Table 1.
Maintenance of cultures
The
cultures were maintained on yeast extract-mannitol medium (YEM) agar (Bohlool
and Schmidt, 1970) slants containing 0.05% CaCO3. Antibiotic resistant mutants and
transconjugants were maintained on tryptone-yeast extract medium (TY) agar
(Beringer, 1974) slants or plates with the appropriate antibiotics (Sigma
Chemical Co., St. Louis, Mo.).
Chloramphenicol was used at 20 ug ml-1, kanamycin at 50 ug ml-1,
streptomycin at 200 ug ml-1, and rifampicin at 20 ug ml-l.
Assessment of culture purity and
identification of strains
The
purity of the cultures was assessed by streaking on YEM agar plates with 0.25
mg 1-1 brom thymol blue. The
plates were examined for colony morphology and acid production.
The
identity of the strains was determined by immunofluorescence (Schmidt et al.,
1968), by intrinsic antibiotic resistance patterns (Josey et al., 1979;
Kingsley and Bohlool, 1983), and by phage typing (Adams, 1959; Dowdle,
1985). The plasmid profile of each
strain was also used to differentate one strain from another. A combination of these methods was used to
check the identity of the isolates from mutation and mating experiments and the
reisolates from plant tests.
|
|
Immunofluorescence
Fluorescent
antibodies (FAs) specific for R. fredii USDA 205 and HH 003 and R.
leguminosarum 6015 (prepared by M. Sadowsky and B.B. Bohlool) were used
to identify these strains. Smears were
made from broth cultures or macerated nodule suspensions, which in the case of R.
fredii, were boiled in a water bath for 30 min to enhance the reaction
of the FA. Gelatine-rhodamine isothiocyanate was used to suppress nonspecific
adsorption (Bohlool and Schmidt, 1968).
The smears were stained with the FAs, and incubated in a moist chamber
for 30 min. The excess stain was washed
off with phosphate buffered saline (PBS), pH 7.2, and the slides soaked in PBS
for 20 min. The slides were rinsed
twice with distilled water, air dried, and observed with a Zeiss standard
microscope equipped for epifluorescence microscopy with a fluorescein
isothiocyanate filter, and a Zeiss condenser with phase objectives. Cross-reactios of FA-stained smears were
quantitated by assessment of the degree of fluorescence from 0 to 4+.
Intrinsic
antibiotic resistance
Filter-sterilized
solutions of antibiotics were added to melted YEM agar which had been cooled to
50oC, to give the following final concentrations (ug ml-1):
chloramphenicol (Chl) 12.0, 25.0; kanamycin sulfate (Kan) 10.0; nalidixic acid
(Nal) 10.0; neomycin sulfate (Neo) 2.5; polymixin B sulfate (Pol) 20.0;
rifampicin (Rif) 1.0, 6.0; streptomycin (Str) 2.5, 10.0; tetracycline hydrochloride
(Tet) 1.0; and vancomycin hydrochloride (Van) 1.5, 5.0. The antibiotic stock solutions were prepared
at a concentration of 10 mg ml-1 in distilled water except Chl (10
mg ml-1 in 95% ethanol), Nal (10 mci ml-1 in 1N NaOH),
and Rif (10 mg ml-1 in methanol).
YEM agar plates without antibiotics served as controls.
To
inoculate several strains on the surface of agar plates simultaneously, 200-ul
volumes of 24-hour old YEM broth cultures were transferred to the wells of a
sterile Falcon 96-well tissue culture plate and then inoculated onto the plates
with a sterile multiple inoculator. Duplicate plates were incubated at 28oC
for seven days and isolates showing growth were scored as positive for
antibiotic resistance.
Phage
typing
Assays
on phage susceptibility were cone on R. fredii HH strains using
the standard agar overlay method (Adams, 1959; Dowdle, 1985). Vincent's defined
agar medium (Vincent, 1970) with 1.0% agar was used as the basal layer, and the
same medium with 0.42% agar was used as the top layer. Cells were grown on YEM broth to
mid-exponential phase and 200 ul of the culture was added to the molten top
agar, spread onto the basal agar layer, and allowed to solidify. The wells of a sterile tissue culture plate
were filled with 200 ul each of phage stock solutions (isolated by S. Dowdle
from soybean-growing regions of China with indigenous populations of R. fredii),
and a flame-sterilized multiple inoculator was used to transfer the phages to
the plate. Phages were not transferred
to the control plates. Duplicate plates
were incubated at 28oC and observed for lysis after 48 h.
Plasmid profile analysis
A
direct lysis procedure was used to isolate plasmid DNA. Plasmid profiles of parent strains and
transconjugants were obtained using a modification of the Eckhardt (1978)
in-well lysis and agarose gel electrophoresis technique described by Rosenberg,
et al. (1982) and M. Kingsley (personal communication).
Cells
were grown overnight in TY, YEM, or peptone (PA) (Hirsch, et al., 1980). Depending on the strain, 200- to 300-ul
volumes of the broth cultures were centrifuged in 1.5 ml plastic Eppendorf
tubes to harvest the cells. As a
preliminary step, the cells were washed with a dilute detergent solution of TEN
buffer (0.05M Tris-base, 0.02M EDTA, 0.02M NaCl) containing 0.1% Sarkosyl
(Schwinghamer, 1980). The pellet was
loosened by vigorous blending in a Vortex mixer, and 40 ul of sphereoplasting
solution was added. This solution
contained 7,500 ug ml-l lysozyme (Sigma), 30 U ml-1 RNAse
I (Worthington Diagnostics, Freehold, N.J.), 20% Ficoll 400 (Sigma), 0.05%
bromo-phenol blue in TEB buffer (89mM Tris-base, 8.9mM EDTA, 89mM boric
acid). The cell suspension was then
layered beneath 200 ul TEB in a well of a 3-mm thick vertical gel. The 0.7% agarose (low mr; Bio-Rad
Laboratories, Richmond, Calif.) gel was prepared by melting the agarose in TEB
in the microwave oven, and readjusting for evaporation loss by adding distilled
water.
After:
a 15-minute incubation to allow for lysozyme action, the lysis procedure was
completed by adding an overlay of 0.2% sodium dodecyl sulfate (SDS) solution in
TEB with 10% Ficoll 400. This was
followed by a second overlay solution of 0.2% SDS solution in TEB with 5% Ficoll
400. The wells were sealed with molten agarose, and electrophoresis was done at
8 mA for 2 h. The current was then
increased to 40 mA and electrophoresis was continued for 5 h. After staining with ethidium bromide (5 ug
ml-1), the plasmid DNA was visualize) with a UV transilluminator,
then photographed using Polaroid 107C film and UV and yellow (Hoya, K2)
filters.
The
approximate molecular weights of the plasmids were determined from their
relative mobilities in agarose gels by using the 130-Mdal pJB5JI and plasmids
of R. leguminosarun (Hirsch et al., 1980), R. trifolii T-12
(Rosenberg et al.,1982), and R. meliloti L5-30 (Rosenberg et al.,
1982) as molecular weight markers.
Development of mutants
Plasmid-cured
mutants
Plasmid
curing was done by acridine orange treatment, by heat treatment, or by a
combination of both.
The
acridine orange treatment was done according to the procedure of Zurkowski et
al. (1973). Overnight YEM broth
cultures were used to inoculate fresh broth containing 1, 3, 5, 10, 15, and 20
ug ml-1 acridine orange (Sigma).
The cultures were incubated in the dark at 25oC on a shaker
for six days. After incubation, the
cultures were diluted in 0.09% (w/v) NaCl and plated on minimal medium (MM)
agar (Vincent, 1970) to exclude auxotrophs.
The resulting colonies were transferred to MM broth to eliminate the
possibility of cross-feeding, then streaked once more on MM plates. Single colonies were picked and their DNA
was observed on agarose gels for changes in their plasmid profiles.
The
procedure of Zurkowski and Lorkiewicz (1978) was used for heat curing. Overnight YEM broth cultures were inoculated
into PA broth. The cultures were
incubated in 35, 37, 40, and 45oC water baths, and given daily
5-second blending in a Vortex mixer for aeration. The cultures were transferred to fresh broth at weekly
intervals. After heat treatment, the
cultures were plated on YEM agar and the plasmid profiles of single colony
isolates were observed.
In
cases where plasmid curing was difficult, the strains were given a combination
of acridine orange and elevated temperature simultaneously.
Antibiotic
resistant mutants
Spontaneous
antibiotic resistant mutants were obtained by growing the cells initially in 50
ml of YEM broth containing 10 ug ml-1 Chl. To boost the level of resistance, the cells were harvested by
centrifugation and transferred to fresh broth with increasing antibiotic
concentrations. The cultures growing in
broth containing 20 ug ml-1 Chl were then streaked on TY plates with
the same antibiotic concentrations.
Bacterial matings
The
procedure for bacterial conjugation was that described by Buchanan-Wollaston et
al. (1980). Crosses were done by
suspending 1 ml of cultures grown overnight that contained approximately 108
cells in sterile saline. The recipient
and the donor cells were filtered successively onto a 0.45-um pore size
Millipore filter (Millipore Corp., Bedford, Mass.). The filters were incubated on TY plates at 28oC
overnight. The cells were collected
from the filter by placing it in an empty sterile Petri dish, adding 5 ml of
sterile saline, and suspending the cells with the aid of a sterile stir
bar. The cell suspensions were serially
diluted and plated on selective and nonselective media.
To
select for R. fredii transconjugants which received the pJB5JI
kanamycin marker, and to counterselect against the donor R. leguminosarum
6015 (pJB5JI), TY medium, supplemented with 50 ug ml-1 Kan and 20 ug
ml-1 Chl, was used. In
backcrosses to the Nod- mutant R. leguminosarum 6015,
TY medium, supplemented with 50 ug ml-l Kan, 20 ug ml-1
Rif, and 200 ug ml-1 Str, was used for selection.
The
same procedure was used on the donor and recipient cultures separately to serve
as controls. The transfer frequency of
each cross was calculated as the number of transconjugants per total number of
recipients. Viable cell counts were made from selective plates using the Miles
and Misra drop-plate count method (Vincent, 1970). R. leguminosarum 6015 (pJB5JI) was counted from a
TY plate containing 50 ug ml-1 Kan while R. fredii was
counted from a TY plate containing 20 ug ml-1 Chl. Viable cells counts were also made from
nonselective TY plates.
Plant infection tests
Plant
culture
Nodulation
tests for soybeans and peas were done in modified Leonard jar assemblies
(Leonard, 1943). The growth assembly
was that used by Sadowsky (1983) and consisted of a 250-ml wide mouth
Erlenmeyer flask which served as the growth vessel and a 500-ml wide mouth
screw-cap bottle which served as the nutrient reservoir. The two components were glued together and
an absorbent cotton wick between the two facilitated the transfer of nutrient
solution from the lower nutrient reservoir into the plant growth vessel
above. The nutrient reservoir was
filled with quarter strength nitrogen-free medium by Hoagland and Arron
(1938). The growth vessel was filled
with a 3:1 (v/v) mixture of washed vermiculite and perlite and then moistened
with the nutrient solution. The growth assembly
was covered with aluminum foil and autoclaved for 45 min (121oC, 15
psi).
Pea,
(Pisum sativum cv. Wisconsin Perfection) and soybean (Glycine
max cv. Peking and Williams) seeds were surface-sterilized by immersion
in a 4.0% (w/v) calcium hypochlorite solution for 20 min, followed by five
washings with sterile distilled water.
The seeds were allowed to imbibe for four h, and then three seeds were
transferred aseptically to each plant growth vessel. The seeds were sown two clays apart to allow five days for the
peas and three days for the soybeans to germinate and grow before inoculation. The seedlings were then thinned to one per
growth vessel, and were inoculated with 1 ml aliquots of 24-hour old YEM broth
cultures. The growth vessels were topped
with 3 cm of silicone-coated sand (5 kg sand:100 ml silicone, Bondex International
Inc., St. Louis, Mo.) to prevent cross-contamination. The plants were incubated in a plant growth chamber at: 25oC
with a daylength of 16 h. In all plant
tests, uninoculated plants and plants inoculated with reference and parent
strains were included for comparison.
Parameters
for symbiotic effectiveness
The
plants were harvested four weeks after sowing and examined for the presence of
nodules (Nod+) and nitrogen fixation (Fix+). The nodulating phenotype was based on gross
nodule development while nitrogen fixation was judged according to plant color,
shoot dry weight, and nitrogenase activity. Plant and nodule dry weights were
determined after drying at 75oC for 48 h. The acetylene reduction
assay (Hardy et al., 1968) was used as an index of nitrogenase activity. The assay was done at 25oC by
incubating excised roots in 60 ml serum bottles with a 5% (v/v) acetylene in
air atmosphere for 30 min. Gas samples
were analyzed for ethylene with a Bendix 2500 gas chromatograph equipped with a
Porapak-T column (80-100 mesh), using a running temperature of 70oC
and a carrier gas flow rate of 50 ml mm-1. The nitrogenase activity was expressed as micromoles of ethylene
formed per hour per plant.
A
one-way analysis of variance was used to determine if the introduction of the
plasmid had a significant effect on the symbiotic performance of the R. fredii
transconjugants. The ANOVA procedure
was done using the SAS statistical package at the University of Hawaii,
Honolulu, HI.
Strain
reisolation
At
the end of the plant tests, the rhizobia were reisolated from root nodules and
their identity was checked to confirm that the root nodules had been formed by
the appropriate strains. Nodules were
excised from the roots and surface-sterilized for 5 s in 75% ethanol, followed
by a 5-minute immersion in 4.0% calcium hypochlorite. After five washings in sterile distilled water, the nodules were
macerated in sterile saline and streaked for isolation into YEM agar plates
containing 0.25 mg 1-1 brom thymol blue.
Plasmid
stability
The
stability of the introduced plasmid was determined by passing the
transconjugants through two plantings of the host plants. The isolates were used to inoculate a second
batch of host plants and the strains recovered after each passage were examined
for the presence of the kanamycin marker by antibiotic testing and the presence
of the plasmid by agarose gel electrophoresis.
CHAPTER
3
RESULTS
Four
R. fredii strains that effectively nodulate both 'Peking' and
'Williams' soybeans, and the type strain, USDA 205, which forms effective
nodules on 'Peking' but ineffective nodules on 'Williams' were the subject of
this study. The strains of R. fredii
and R. leguminosarum that were used were characterized in terms
of acid production, FA reaction, IAR pattern, and phage susceptibility.
All
of the strains produced acid, turning the YEM agar medium which contained brom
thymol blue yellow after three days of growth.
None of the R. fredii strains shared cross reactive
antigens with R. leguminosarum 6015 while HH 003 cross reacted
with FA 205 (Table 2). USDA 205 and HH
003 also exhibited the same IAR pattern (Table 3) and HH 103 could be
distinguished from HH 102 and HH 303 by its sensitivity to 10 ug ml-1
of kanamycin. The R. leguminosarum
strains and USDA 205 were not susceptible to any of the phages used, while each
of the HH strains were susceptible to one phage each, with HH 102 and HH 103
sharing the same phage-typing pattern (Table 4).
Plasmid profiles
Indigenous
high-molecular weight plasmids were found in all of the R. fredii
strains examined. In all strains except
HH 303, plasmids that are greater than 300 Mdal in size (megaplasmids) were
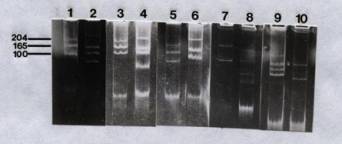
also observed. Figure 1 shows the
relative electrophoretic mobilities of plasmids from USDA 205 and the HH
strains 003, 102, 103, and 303, together with the reference plasmids used to
estimate plasmid sizes. Each of the
strains had a unique plasmid profile.
They contained at least two plasmids, four in USDA 205, HH 003 and HH
103, and three in HH 102.
|
|
|
29 |
|
|
|
|
Figure 1.
Agarose gel electrophoresis of plasmid DNA from R. fredii. Lane: 1 and 5, R. leguminosarum 6015; 2, USDA 205;
3, R. trifolii T-12; 4, HH 003; 6, HH 102; 7, R. meliloti
L5-30; 8, HH 103; 9, R. leguminosarum 6015 (pJB5JI); and 10, HH
303. The numbers refer to the sizes of
the plasmids in Mdal. (Picture of HH
003 and reference plasmid is courtesy of M. Kingsley)
|
|
Isolation of plasmid cured-mutants
In
order to determine whether genes for symbiotic functions are carried on the
plasmids of the R. fredii strains, they were subjected to
treatments designed to eliminate the plasmids.
The
plasmid-curing experiments resulted in a change in the plasmid profiles of some
of the strains. The plasmid cured
strains were designated according to the treatments used to produce them. USDA 205-A05
was isolated after exposure to 5 ug ml-1 AO, HH 003-1C40 and HH 003-2C40 after one and two week cycles of growth at 40oC
respectively, and HH 102-AO2T37 after
exposure to 2 ug ml-1 AO at 37oC. As shown in Figure 2, USDA 205-AO5 had lost
its third largest plasmid, HH 003-2C40,
its second largest plasmid, and HH
102-A02T37, its third largest one.
HH003-1C40 lost its second and
third plasmids but gained one that showed a lower electrophoretic mobility,
possibly as a result of cointegration of two plasmids. There were no differences between the cured
strains and their parents with regard to their growth on YEM, FA reaction, IAR
pattern, and phage susceptibility (data not shown).
Figure 2. Plasmid
profiles of R. fredii cured mutants and parent strains. Lane: 1, USDA 205-A05; 2, USDA 205; 3, HH
003; 4, HH 003-1C40; 5, HH 003-2C40; 6, HH 102; and 7, HH 102-A02T37.
|
|
|
|
The
plasmid-cured strains were tested on host plants and their symbiotic phenotypes
are presented in Table 5. Two of the
cured strains (USDA 205-A05 and HH 003-2C40) failed to nodulate soybeans while
HH 003-1C40 and HH 102-A02T37 formed effective nodules like the parent strains.
HH
103 and HH 303 both failed to develop cured mutants, even after subjecting them
to a combined AO-high temperature treatment for five weeks.
Transfer, expression, and maintenance of pSym
plasmid
The
R. fredii strains were used as recipients of pJB5JI, a 130-Mdal
derivative of the conjugative and bacteriocinogenic plasmid pRLIJI from R.
leguminosarum into which the transposon Tn5 has been introduced
(Beringer et al. 1978). This plasmid
encodes five recognizable functions: the host specificity for pea, the
nodulation functions, the N2-fixing ability, the transfer function
(tra), and kanamycin resistance (Johnston et al., 1978). The nodulation
functions include root adhesion (Roa+), hair curling (Hac+),
and nodule initiation (Noi+) (Rolfe et al., 1983). Kanamycin resistance is controlled by Tn5
that was inserted in the genes for medium bacteriocin production (Mbp::Tn5),
and is used as a marker to monitor the presence of the plasmid. Hence,
selection for the inheritance of pJB5JI by the recipients in the conjugation
experiments was for kanamycin resistance.
To
determine if the R. fredii strains could be used as recipients of
pJB5JI, they were tested for resistance to the kanamycin levels used in the
mating experiments. Only HH 102 and HH
303 which showed natural resistance to a low level of kanamycin in their IAR
patterns, exhibited growth beyond 10 ug ml-l of kanamycin (Table
6). Resistant colonies of HH 003 were
present at 50 ug ml-1 of kanamycin, and precluded the use of the
strain in the mating experiments.
|
|
Transfer
of pJB5JI
The
ability of R. fredii strains to receive, express, and maintain
pJB5JI from R. leguminosarum 6015 was studied by using parent and
plasmid-cured strains in bacterial conjugation experiments.
The
transfer of the Tn5-encoded kanamycin resistance marker occurred at low
frequencies (Table 7), with transfer to HH 102 being the highest at 5 X 10-5,
and transfer to USDA 205, the lowest at 2 X 10-8. Parent strains and their plasmid-cured
derivatives had similar transfer frequencies, with USDA 205 and USDA 205-A05
having a frequency of 10-8 and HH 003 and HH 003-2C40 having a
frequency of 10-7.
Analysis
of the plasmid content of the R. fredii transconjugants were made
to determine whether their Kanr phenotype was due to an intact
pJB5JI, or to spontaneous mutation or transposition of the Tn5 into the
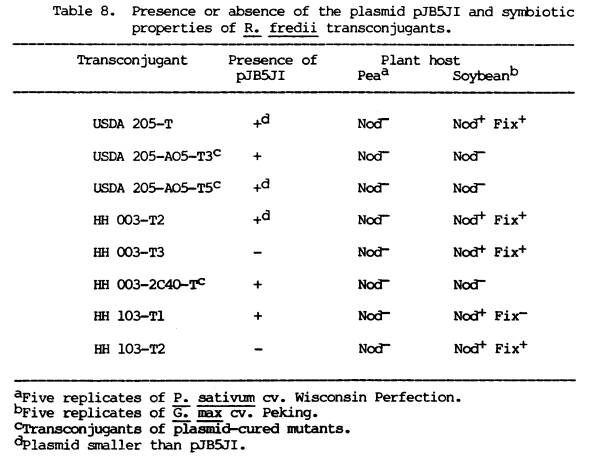
chromosome or resident plasmids. Table
8 is a summary of the plasmid content of the R. fredii
transconjugants determined from agarose gels, showing the presence or absence
of p7B5JI. Three of the transconjugants
had acquired a new plasmid band with the same electrophoretic mobility as
pJB5JI, three displayed a plasmid band that indicated a plasmid smaller than
PJB5JI, and two were indistinguishable from the corresponding recipient
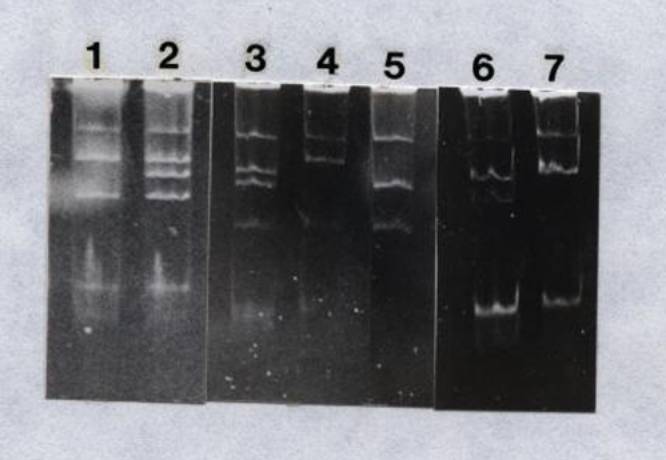
strains. Figure 3 presents the plasmid
profiles of the USDA 205-AO5 transconjugants, and for reference, those of the
wild type USDA 205, the donor R. leguminosarum 6015 (pJB5JI), and
the recipient strain USDA 205-AO5.
|
|
|
|
|
|
Table
8 also outlines the symbiotic properties of the R. fredii
transconjugants. In order to determine
the effect of the introduced plasmid genes, plant hosts were used to screen the
transconjugants for gene expression.
The symbiotic phenotypes of the transconjugants were examined by
inoculation of soybean plants, the normal host of R. fredii and
pea plants, the host specified by the pJB5JI-encoded genes. None of the R. fredii
transconjugants formed nodules on peas.
The transconjugants of the plasmid-cured strains still did not nodulate
soybeans but those of the parent strains did.
The
presence of pJB5JI in the transconjugants was also confirmed by backcrossing
them to the Nod- R. leguminosarum 6015. The R. fredii transconjugants
were able to transfer the pJB5JI kanamycin marker at relatively higher frequencies, ranging from 10-4
to 10-6 (Table 9). The
transconjugants of USDA 205 and HH 003, and those of their plasmid-cured
derivatives showed similar transfer frequencies of 10-4.
The
R. leguminosarum 6015 transconjugants from the backcrosses had a
variety of plasmid profiles (Figure 4).
Based on their plasmid make-up, the 6015 transconjugants can be grouped
into two categories: those that received an additional band that had similar
mobility as pJB5JI and those that did
not. Within these two groups,
additional variation can be seen. There
are four kinds of plasmid profiles in the first group: first, those that
resemble 6015 (pJB5JI) (lane 3); second,
those that show an additional high mobility plasmid band that comigrated with
the linear chromosomal fragments (lane 4); third, those that show two
additional plasmid bands (lane 5); and fourth, those that are missing the
smallest plasmid of 6015 (lane
6). The group that did not show the pJB5JI plasmid band are of two kinds: those
that resembled the recipient strain 6015 (lane
7), and those that have lost the smallest 6015
plasmid (lane 8).
Figure 3.
Plasmid profiles of R. fredii USDA 205-A05 transconjugants
and reference strains. Lane: 1, USDA
205-A05-T3; 2, USDA 205-A05-T5; 3, USDA 205-A05; 4, USDA 205; and 5, R. leguminosarum
6015 (pJB5JI). Arrow points at plasmid
pJB5JI.
|
|
|
|
|
|
Figure 4.
Plasmid profiles of R. leguminosarum 6015 and its
transconjugants. Lane: 1, 6015
(pJB5JI); 2, 6015; 3, representative profile of T2 and T7A; 4, T7C; 5, T7B; 6,
T5A; 7, representative profile of T1, T3A, T3B, T4, T6, and T8; and 8, T5C. Arrow points at plasmid pJB5JI.
|
|
|
|
Table
10 presents a summary of the pJB5JI donor,
plasmid make-up and symbiotic properties of the R. leguminosarum
transconjugants resulting from the backcrosses. None of the transconjugants nodulated soybean plants, and those
that did not receive pJB5JI failed to
nodulate pea plants as well. The
transconjugants with pJB5JI were
nodulation competent and formed either effective or ineffective nodules on
peas.
Symbiotic
effectiveness of transconjugants
The
effect of pJB5JI on the nitrogen
fixation ability of the nodulation competent R. fredii
transconjugants was assessed by comparing their symbiotic performance on
soybean plants with those of the wild type parents. As Table 11 shows, the transconjugants exhibited various degrees
of effectiveness. While the HH 003
trans-conjugants were identical to the parent strain in all measured parameters
of symbiotic effectiveness, USDA 205-T differed significantly from its parent
strain in terms of the amount of ethylene produced and shoot dry weight. HH 103-Tl was ineffective in nitrogen
fixation, the plants having very small white nodules and the leaves appearing
yellow and chlorotic like the uninoculated plants. HH 103-T2 formed effective nodules but differed significantly
from the parent strain in the amount of ethylene produced, nodule dry weight
and shoot weight.
|
|
|
|
On
the second passage through pea and soybean plants, the reisolated
transconjugants still did not nodulate peas, but retained their ability to
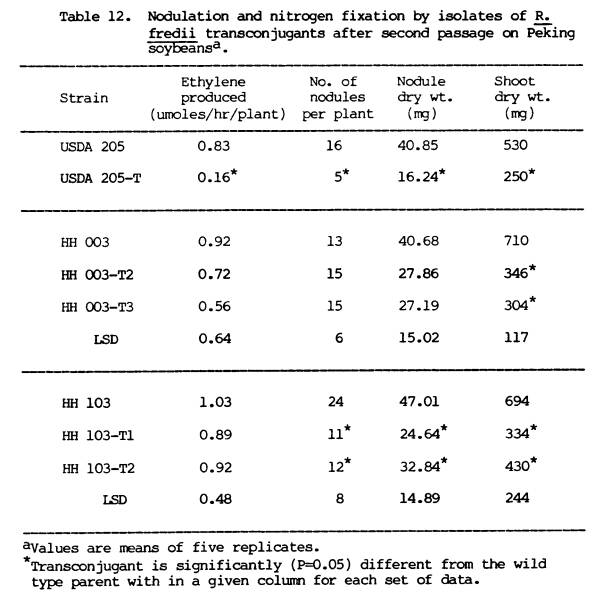
nodulate soybeans. Table 12 presents
the symbiotic performance of the R. fredii transconjugants on
second passage on soybean plants. USDA
205-AO5-T still differed significantly from its parent strain, but this time in
all the observed parameters. The two
transconjugants of HH 003 were as effective as the parent, except in terms of
shoot dry weight. HH 103-T1 formed effective
nodules but differed significantly from the parent in nodule dry weight and
shoot dry weight.
The
reisolated transconjugants retained their antibiotic markers, and there were no
detectable changes in their plasmid profiles (data not shown).
|
|
CHAPTER 4
DISCUSSION
This
study confirms and extends earlier studies that showed the presence of large
indigenous plasmids in R. fredii and their involvement in
determining symbiotic properties.
Each
of the R. fredii strains examined contained high molecular weight
plasmids and all can be differentiated on the basis of their plasmid
profiles. The plasmid profile of the
type strain USDA 205, consists of four plasmid bands and is identical to that
of other studies (Appelbaum et al., 1985a; Broughton et al., 1984; Masterson et
al., 1985). Previous work on this
strain showed three plasmid bands ranging from 57 to 192 Mdal in size
(Masterson et al., 1982; Sadowsky and Bohlool, 1983). By using a plasmid isolation procedure that is more sensitive to
large covalently closed circular DNA molecules,
a fourth band of lower electrophoretic mobility that is estimated to be greater
than 300 Mdal, was detected. Heron and
Pueppke (1984) reported a 35-Mdal plasmid in addition to these four, but it was
not seen consistently in this study and could not easily be distinguished from
the band of linear chromosomal fragments.
The
presence of megaplasmids in USDA 205 and the HH strains (except HH 303), and in
other strains as well (Appelbaum et al., 1985a; Broughton et al., 1984)
suggests that it may be a common feature in R. fredii as it is in
R. meliloti, R. trifolii, and R. leguminosarum
(Denarie et al., 1981a).
Various
intercalating agents, particularly acridines, inhibit plasmid replication
without inhibiting chromosomal replication.
After exposure to acridine orange, USDA 205-A05 lost its third largest
plasmid, and along with it, the ability to nodulate its soybean host. This indicated the involvement of the
missing plasmid in the symbiotic functions.
Physical
studies of this 112-Mdal plasmid, designated pRjaPRC205b by Masterson et al.
(1982), show the presence of genes for nodulation and nitrogen fixation. The presence of nod genes was
indicated by hybridization with pRmSL26, a cosmid clone that contains a 20-kb
insert of R. meliloti DNA having several nod genes (Long
et al., 1982; Appelbaum et al., 1985a), and with pRs23, a plasmid containing nod
sequences from a Rhizobium strain that nodulates tropical legumes and
the nonlegume Parasponia (Djordjevic et al., 1985a). The genes for nitrogen fixation were located
by hybridization with pRmR2, a recombinant R. meliloti plasmid
with a 3.9-kb EcoRI fragment homologous to the nifD and nifH
genes in K. pneumoniae (Masterson et al., 1982; Ruvkun and
Ausubel, 1980). Multiple copies of the nod
and nif sequences on this 112-Mdal plasmid was reported by Prakash and
Atherly (1984).
The
nod genes are duplicated in two other plasmids in USDA 205. Appelbaum et
al. (1985a) reported that the 192-Mdal and 57-Mdal plasmids also carry nod
genes, since they also hybridized with pRmSL26. Sadowsky and Bohlool (1983) have shown that elimination of the
192-Mdal plasmid resulted in the loss of nodulation ability. From these studies, it appears that in USDA
205, the remaining gene copies after plasmid curing cannot compensate for the
loss of the plasmid, and leads to the loss of the symbiotic function.
However,
in studies of another R. fredii strain, USDA 206, curing of a
197-Mdal plasmid carrying both nif and nod genes did not result
in the loss of any symbiotic functions, but to a reduction of nodulation and
nitrogen fixation performance (Mathis et al., 1985). Barbour et al. (1985) suggested the presence of other functional nif
genes in the chromosome and in another plasmid, but the duplicate nod
genes that are responsible for the retention of the nodulation function were
not located.
Prolonged
exposure to elevated temperatures is another factor that results in the loss of
plasmids, and it was postulated that this was due to its effect on the
temperature sensitive plasmid replication process (Zurkowski, 1982). After seven days of heat treatment, HH
003-1C40 had two missing plasmid bands, but gained a new band of lower
mobility. The large size of the new
plasmid indicates that it may be a cointegrate of two plasmids. The formation of effective nodules by HH
003-1C40 on soybeans shows that functional nod genes are still present.
HH
003-2C40 lost its second largest plasmid after a two-week cycle of growth at
high temperature. The loss of this
large plasmid, which comigrated with the 460-Mdal megaplasmid of R. trifolii
T-12 (Zurkowski and Lorkiewicz, 1979) resulted in the loss of the ability to
nodulate soybeans. This points to a megaplasmid as the possible location of
symbiotic genes. This is unusual because, in R. fredii,
nodulation and nitrogen fixation genes are generally known to be carried on
150-200 Mdal plasmids (Broughton et al., 1984; Masterson et al., 1982). In studies done by Appelbaum et al. (1985a),
hybridization experiments using nif and nod probes on R. fredii
plasmids failed to show homology with the megaplasmids, but did so with the
medium-sized plasmids in eight strains and with the chromosome in two strains.
The
third largest plasmid of HH 102-A02T37 appears to be a cryptic plasmid. Its loss did not affect the symbiotic
performance of the strain, and its functions are unknown.
Like
previous work done by Sadowsky and Bohlool (1985), the results of the
plasmid-curing experiments correlate the absence of nodulation ability with the
loss of plasmids. However, the nature
of the changes in the nodulation process effected by the plasmid loss remains
to be defined.
At
least one of the plasmids of HH 103 is transmissible to R. leguminosarum
6015. The transfer of pJB5JI from the
transconjugant HH 103-T1 to 6015 resulted in the appearance of two new plasmid
bands, in addition to the pJB5JI plasmid band (Figure 4, lane 5). The larger of the two new plasmids appears
to be one of the plasmids of HH 103-T1 since it showed similar electrophoretic
mobility. There is no plasmid band in
HH 103-T1 that corresponds to that of the smaller plasmid indicating that this
may not have been previously detected or may represent one of the native
plasmids that suffered a deletion.
The
R. leguminosarum plasmid, pJB5JI, is transmissible between
different strains of R. leguminosarum, R. trifolii,
and R. phaseoli at high frequency, occurring at 10-2
per recipient (Johnston et al., 1978).
In the same study, the establishment of the plasmid-linked markers in R.
meliloti was lower at 10-4 per recipient. The R. fredii strains in this
study are able to conjugate with R. leguminosarum and act as
recipients of pJB5JI at low frequencies (10-5 to 10-8). Studies of other strains showed higher
frequencies ranging from 10-2 to 10-5 (Ruiz-Sainz et al.,
1984; Sadowsky, 1983), except in the case of USDA 205. The transfer frequency of USDA 205 in this
study was slightly higher than that of Sadowsky (1983), which showed a low 10-9
transfer frequency of the kanamycin marker.
Transfer of pJB5JI from the R. fredii transconjugants back
to R. leguminosarum occurred at frequencies that are similar to
those obtained by Sadowsky (1983) at 10-4 per recipient.
Whether
the R. fredii recipient had its normal complement of plasmids or
not, transfer frequency remained the same for both the parent and the plasmid-cured
derivative. This indicates the lack of
an entry exclusion function on the missing plasmids.
The
inability of the kanamycin resistant R. fredii transconjugants to
form nodules on peas may have a number of causes. First, the transconjugants may not have received all the pJB5JI
genes necessary for nodulation. This is
indicated by the fact that some of the plasmid profiles show a plasmid smaller
than pJB5JI or none at all. It is
therefore possible that only a part of the Tn5-carrying plasmid was
transferred. This is confirmed by the
results of the backcrosses to the Nod- R. leguminosarum
6015 wherein only some of the 6015 transconjugants regained the ability to form
effective nodules on peas. The results
also indicate that there may have been varying sizes of the deletions suffered
during the transfer, some of which are too small to be discernible in the
plasmid profiles. This is evidenced by
the range of phenotypes exhibited by the 6015 transconjugants.
Secondly,
although the R. leguminosarum genes were transferred with the
pJB5JI, they were not expressed in the R. fredii genetic
background. Earlier work by Sadowsky
and Bohlool (1985) showed the lack of function of pJB5JI genes in R. fredii
strains.
The
presence of the plasmid pJB5JI in the plasmid-cured transconjugants did not
enable them to nodulate peas, nor did it restore their ability to nodulate
soybeans. The R. fredii
205-A05-T transconjugant received intact nod genes as confirmed by the
Nod+ Fix- phenotype of the 6015 transconjugant that
resulted from the backcross, yet the introduction of the plasmid did not
replace the genetic information needed by the recipient to nodulate its normal
host. In one study, pJB5JI
transconjugants of a cured Nod- R. trifolii strain
formed effective nodules on peas, and later formed ineffective nodules on
clover (Sadowsky and Bohlool, 1985).
This is consistent with the known taxonomic relationship of the species,
considering the high degree of genetic homology between R. trifolii
and R. leguminosarum. In
the case of the more distantly related R. meliloti, Fisher et al.
(1985) reported that several nodulation genes are functionally replaceable by a
cloned nod gene DNA fragment from R. trifolii and that R.
meliloti clones like-wise complement two R. trifolii Nod-
mutants. The introduction of pJB5JI to
a Nod- R. meliloti that suffered a deletion during
heat treatment enabled the transconjugant to form ineffective nodules on
alfalfa (Banflavi et al., 1981). In the other Nod- mutants tested,
no restoration of the nodulation ability was found.
In
this study, the failure of the plasmid-cured transconjugants to regain their
nodulation ability with pJB5JI, may indicate that the symbiotic genes lost
through the curing process were too substantial, and thus, not replaceable by
the common nod genes. It may
also indicate that R. fredii represents a genetically separate
class of fast-growing rhizobia and the nod genes are too evolutionarily
diverged to allow complementation.
As
in the case of the transconjugants of the plasmid-cured derivatives, there was
no observable expression of the PJB5JI host range genes by the transconjugants
of the parent strains. Using other R.
fredii strains, Sadowsky and Bohlool (1985) reported similar results,
but Ruiz-Sainz et al. (1984) observed swellings on pea roots which
microscopically resembled the early stages of nodule development.
The
presence of pJB5JI elicited different effects on the symbiotic performance of
the Nod+ R. fredii transconjugants on soybeans. Some transconjugants formed effective
nodules comparable to the wild-type strains, some showed a reduction in their N2-fixing
effectiveness while one transconjugant strain did not fix nitrogen at all.
Johnston
et al. (1978) suggested that the presence of genetic information for the
nodulation of host plants of different cross inoculation groups in the same Rhizobium
strain might cause the strain to be impaired in its nodulation on either
host. It is postulated that this
phenomenon might be due to some form of functional interference occurring
between the two plasmids specifying nodulation for different host legumes. Beynon et al. (1980) observed that
inoculation with a strain that contained both the R. phaseoli nodulation
plasmid and pJB5JI resulted in the recovery of clones from pea nodules that
either suffered a subtantial deletion in the R. phaseoli
nodulation plasmid or else lost it completely.
This suggested that genetic information on this plasmid had to be
eliminated spontaneously before nodulation specified by the R. leguminosarum
plasmid pJB5JI could proceed normally.
According to Nadler (1984), genes in the symbiotic region of pJB5JI
inhibited nodulation of soybeans by R. fredii strains. USDA 205 (pJB5JI) transconjugants induced
nodules poorly on roots of Peking soybeans whereas transconjugants containing a
plasmid derived from pJB5JI by a 20-Mdal deletion removing the symbiosis genes
induced nodules similar to those of the parental USDA 205.
In
this study, the symbiotic properties of the HH 103 transconjugants (Table 8)
show the plasmid effect. HH 103-T1, which received the intact pJB5JI, was Fix-,
while HH 103-T2, which has kanamycin resistance but no pJB5JI, was Fix+. These results show that there is functional
interference between the symbiotic genes, and further indicate that the
impairment of the symbiotic process may extend to N2-fixing
effectiveness as well.
Transconjugants
isolated from the soybean nodules retained their antibiotic resistance markers,
but showed changes in their symbiotic properties when used in a second cycle of
plant inoculation. This may indicate
that gene expression has been modified during nodule development. The observation that a Fix-
transconjugant in the first cycle of inoculation regained its effectiveness in
the second cycle points to the relative instability of pJB5JI. The variable
stability of pJB5JI in different
strains of R. trifolii was reported by Sadowsky and Bohlool
(1985). In their study, reisolates of
transconjugants from pea and clover nodules exhibited a range of symbiotic
properties. Similar behavior of the Fix+
phenotype of wild-type R. trifolii strains carrying pJB5JI was reported previously by Djordjevic
et al. (1982).
The
reisolated transconjugants retained their antibiotic resistance markers, and
showed the same plasmid profiles, suggesting that the plasmid is maintained
without gross alterations. However,
since there were differences of symbiotic effectiveness of the transconjugants
after passage through the host plants, there may have been alterations in the
plasmid that were not detected in the agarose gels.
In
summary, this study shows that high molecular weight plasmids that are involved
in the control of symbiotic functions, together with others whose functions are
not known, are an integral part of the genetic make-up of R. fredii
strains. The loss of symbiotic
functions is correlated with the loss of plasmids. These strains are capable of accepting and maintaining pJB5JI, a pSym plasmid from R. leguminosarum. Although pJB5JI
plasmid genes are not expressed, they appear to affect the expression of
R. fredii symbiotic genes.
LITERATURE CITED
Adams, M.H. 1959. Bacteriophages. Interscience,
New York.
Appelbaum, E.R., E. Johansen, and N.
Chartrain. 1984. Identification of plasmids carrying
symbiotic genes in fast-growing Rhizobium japonicum using DNA
hybridization and Tn5 mutagenesis, p. 670. In:
C. Veeger and W.E. Newton (ed.), Advances in nitrogen fixation research.
Nijhoff/Junk, The Hague.
Appelbaum, E.R., E. Johansen, and N.
Chartrain. 1985. Symbiotic mutants of USDA 191, a
fast-growing Rhizobium that nodulates soybeans. Mol. Gen. Genet. 201: 454-461.
Appelbaum, E.R., T.J. McLoughlin, M.
O'Connell, and N. Chartrain. 1985.
Expression of symbiotic genes of Rhizobium japonicum USDA 191 in
other rhizobia. J. Bacteriol. 163:
385-388.
Banfalvi, Z., V. Sakanyan, C. Koncz, A. Kiss,
I. Dusha, and A. Kondorosi. 1981.
Location of nodulation and nitrogen fixation genes on a high molecular
weight plasmid of R. meliloti.
Mol. Gen. Genet. 184: 318-325.
Berg, D.E., and C.M. Berg. 1983.
The prokaryotic transposable element Tn5. Biotechnology 1: 417-435.
Beringer, J.E. 1974. R factor transfer
in Rhizobium leguminosarum.
J. Gen. Microbiol. 84: 188-198.
Beringer, J.E., J.L. Beynon, A.V. Buchanan-Wollaston,
and A.W.B. Johnston. 1978. Transfer of
the drug resistance transposon Tn5 to Rhizobium. Nature 276: 633-634.
Beringer, J.E., N.J. Brewin, and A.W.B.
Johnston. 1980. The genetic analysis of Rhizobium in
relation to symbiotic nitrogen fixation.
Heredity 45: 161-186.
Berry, J.O., and A.G. Atherly. 1984.
Induced plasmid-genome rearrangements in Rhizobium japonicum. J. Bacteriol. 157: 218-224.
Beynon, J.L., J.E. Beringer, A.V.
Buchanan-Wollaston, P.R. Hirsch, and A.W.B. Johnston. 1980a. The involvement of
plasmids in the determination of host specificity in Rhizobium. Soc. Gen. Microbiol. Quart. 7: 75-76.
Beynon, J.L., J.E. Beringer, and A.W.B.
Johnston. 1980b. Plasmids and host-range in Rhizobium leguminosarum
and Rhizobium phaseoli.
J. Gen. Microbiol. 120: 421-429.
Bohlool, B.B., and E.L. Schmidt. 1968.
Nonspecific staining: its control in immunofluorescence examination of
soil. Science 162: 1012-1014.
Bohlool, B.B., and E.L. Schmidt. 1970.
Imnunofluorescent detection of Rhizobium japonicum in
soils. Soil Sci.110: 229-236.
Bohlool, B.B., and E.L. Schmidt. 1984.
Lectins: a possible basis for specificity in the Rhizobium-legume
root nodule symbiosis. Science 185:
269-271.
Brewin, N.J., J.E. Beringer, A.V.
Buchanan-Wollaston, A.W.B. Johnston, and P.R. Hirsch. 1980a. Transfer of
symbiotic genes with bacteriocinogenic plasmids in Rhizobium leguminosarum. J. Gen. Microbiol. 116: 261-270.
Brewin, N.J., J.E. Beringer, and A.W.B.
Johnston. 1980b. Plasmid-mediated transfer of host-range
specificity between two strains of Rhizobium leguminosarum. J. Gen. Microbiol. 120: 413-420.
Brewin, N.J., T.M. DeJong, D.A. Philips, and
A.W.B. Johnston. 1980c. Co-transfer of determinants for hydrogenase
activity and nodulation ability in Rhizobium leguminosarum. Nature 288: 77-79.
Brewin, N.J., J.L. Beynon, and A.W.B.
Johnston. 1981. The role of Rhizobium plasmids in
host specificity, p. 65-67. In J.M.
Lyon, R.C. Valentine, D.A. Philips, D.W. Rains, and R.C. Huffaker (ed), Genetic
engineering of symbiotic nitrogenfixation.
Plenum Press, New York.
Broughton, W.J., N. Heycke, H. Meyer, and
C.E. Pankhurst. 1984. Plasmid-linked nif and nod genes in
fast-growing rhizobia that nodulate Glycine max, Phosphocarpus
tetragonolobus, and Vigna unguiculata. Proc. Natl. Acad. Sci. 81: 3093-3097.
Buchanan-Wollaston, A.V., J.E. Beringer, N.
Brewin, P.R. Hirsch, and A.W.B. Johnston.
1980. Isolation of symbiotically
defective mutants in Rhizobium leguminosarum by insertion of the
transposon Tn5 into a transmissible plasmid.
Mol. Gen. Genet. 178: 185-190.
Burkhardt, B., and H.J. Burkhardt. 1984.
Visualization and exact molecular weight determination of a Rhizobium
meliloti megaplasmid. J. Mol.
Biol. 175: 213-218.
Casse, F., C. Boucher, J.S. Julliot, M.
Michel, and J. Denarie. 1979. Identification and characterization of large
plasmids in Rhizobium meliloti using agarose gel
electrophoresis. J. Gen. Microbiol.
113: 229-242.
Christensen, A.H., and K.R. Schubert. 1983.
Identification of a Rizobium trifolii plasmid coding for
nitrogen fixation and nodulation genes and its interaction with pJB5JI, a Rhizobium
leguminosarum plasmid. J.
Bacteriol. 156; 592-599.
Clowes, R.C., E.E.M. Moody, and R.H.
Pritchard. 1965. The elimination of extrachromoscmal elements
in thymineless strains of Escherichia coli K12. Genet. Res.,
Camb. 6: 147.
Cole, M.A., and G.H. Elkan. 1973.
Transmissible resistance to penicillin G, neomycin, and choramphenicol
in Rhizobium japonicum.
Antimicrob. Agents Chemother.4: 248-253.
Dazzo, F.B. and D.H. Hubbell. 1975.
Cross-reactive antigens and lectins as determinants of symbiotic
specificity in Rhizobium-clover association. Appl. Env. Microbio1.30: 1017-1033.
Denarie, J., P. Boistard, F. Casse-Delbart,
A.G. Atherly, J.0. Atherly, and P. Russel.
1981a. Indigenous plasmids of Rhizobium,
p. 225-264. In K.L. Giles and A.G.
Atherly (ed.), Biology of the Rhizobiaceae.
AcademicPress, New York.
Denarie, J., C. Rosenberg, P. Boistard, G.
Truchet, and F. Casse-Delbart.
1981b. Plasmid control of
symbiotic properties in Rhizobium meliloti, p. 137-141. In A.H. Gibson and W.E. Newton (ed), Current
perspectives in nitrogen fixation.
Australian Academy of Sciences, Canberra.
Djordjevic, M.A., W. Zurkowski, and B.G. Rolfe. 1982.
Plasmids and stability of symbiotic properties of Rhizobium trifolii. J. Bacteriol. 151: 560-568.
Djordjevic, M.A., W. Zurkowski, J. Shine, and
B.G. Rolfe. 1983. Sym plasmid transfer to various symbiotic
mutants of Rhizobium trifolii, R. leguminosarum,
and R. meliloti. J.
Bacteriol. 156: 1035-045.
Djordjevic, M.A., P.R. Schofield, R.W. Ridge,
N.A. Morrison, B.J. Bassam, J. Plazinski, J.M. Watson, and B. G. Rolfe. 1985a.
Rhizcbium nodulation genes involved in root hair curling (Hac) are
functionally conserved. Plant Mol.
Biol. 4: 147-160.
Djordjevic, M.A., P.R. Schofield, B.G.
Rolfe. 1985b. Tn5 mutagenesis of R. trifolii host-specific
nodulationgenes result in mutants with altered host-range ability. Mol. Gen. Genet. 200: 463-471.
Dowdle, S.F.
1985. The ecology of Rhizobium
japonicum in soybean-rice cropping systems in central China. Ph.D. thesis. University of Hawaii.
Downie, A., G. Hcmbrecher, Q.S. Ma, C.D.
Knight, B. Wells, and A.W.B. Johnston.
1983. Cloned nodulation genes of
Rhizobium leguminosarum determine host-range specificity. Mol. Gen. Genet. 190: 359-365.
Eckhardt, T.
1978. A rapid method for the
identification of plasmid deoxyribonucleic acid in bacteria. Plasmid 1: 584-588.
Fisher, R.F., J.K. Tu, and S.R. Long. 1985.
Conserved nodulation gene in Rhizobium meliloti and Rhizobium
trifolii. Appl. Environ.
Microbiol. 49: 1432-1435.
Gross, D.C., A.K. Vidaver, and R.V.
Klucas. 1979. Plasmids, biological properties and efficacy of nitrogen fixation
in Rhizobium japonicum indigenous to alkaline soils. J. Gen. Micro. 114: 257-266.
Hattri, J., and D.A. Johnson. 1984.
Fast-growing Rhizobium japonicum that effectively
nodulates several commercial Glycine max L. Merrill
cultivars. Appl. Environ. Microbiol. 48:
234-235.
Hardy, R.W.F., R.D. Holsten, E.K. Jackson,
and R.C. Burns. 1968. The acetylene-ethylene assay for N2-fixation:
laboratory and field evaluation. Plant
Physiol. 43: 1185-1207.
Heron, D.S., and S.G. Pueppke. 1984.
Mode of infection, nodulation specificity, and indigenous plasmids of 11
fast-growing Rhizobium japonicum strains. J. Bacteriol. 160: 1061-1066.
Higashi, S.
1967. Transfer of clover
infectivity of Rhizobium trifolii to Rhizobium phaseoli
as mediated by an episomic factor. J. Gen.
Appl. Microbiol. 13:391-403.
Hirsch, P.R.
1979. Plasmid-determined
bacteriocin production by Rhizobium leguminosarum. J. Gen. Micro. 113: 219-228.
Hirsch, P.R., M. van Montagu, A.W.B.
Johnston, N.J. Brewin, and J. Schell. 1980.
Physical identification of bacteriocinogenic, nodulation, and other
plasmids in strains of Rhizobium leguminosarum. J. Gen. Microbiol. 120: 403-412.
Hoagland, D.R., and D.I. Arron. 1938.
The water culture method for growing plants without soil. Calif. Agric. Exp. Stn. Circ. 347.
Hombrecher, G., N.J. Brewin, and A.W.B.
Johnston. 1981. Linkage of genes for nitrogenase and
nodulation ability on plasmids in Rhizobium leguminosarum and R.
phaseoli. Mol. Gen. Genet. 182:
133-136.
Hombrecher G., R. Gotz, N.J. Dibb, J.A.
Downie, A.W.B. Johnston, and N.J. Brewin.
1984. Cloning and mutagenesis of
nodulation genes from Rhizobium leguminosarum T0M, a strain with
extended host range. Mol. Gen. Genet.
194:293-298.
Hooykass, P.J.J., A.A.N. van Brussel, H. den
Dulk-Ras, G.M.S. van Slogteren, and R.A. Schilperoort. 1981.
Sym plasmid of Rhizobium trifolii expressed in different
rhizobial species and Agrobacterium tumefaciens. Nature 291: 351-353.
Johnston, A.W.B., J.L. Beynon, A.V.
Buchanan-Wollaston, S.M. Setchell, P.R. Hirsch, and J.E. Beringer. 1978.
High frequency transfer of nodulating ability between strains and
species of Rhizobium. Nature
276: 634-636.
Johnston, A.W.B., G. Hombrecher, N.J. Brewin,
and M.C. Cooper. 1982. Two transmissible plasmids in Rhizobium
leguminosarum strain 300. J.
Gen. Microbiol. 128: 85-93.
Josey, D.P., J.L. Beynon, A.W.B. Johnston,
and J.E. Beringer. 1979. Strain identification in Rhizobium
using intrinsic antibiotic resistance.
J. Appl. Bacteriol. 46: 343-350.
Keister, D.L. 1975. Acetylene reduction
by pure cultures of rhizobia. J.
Bacteriol. 123: 1265-1268.
Kennedy, C., B. Dreyfus, and J. Brockwell.
1981. Transfer, maintenance, and
expression of P plasmids in strains of cowpea rhizobia. J. Gen. Microbiol. 125: 233-240.
Keyser, H.H., B.B. Bohlool, T.S. Hu, and D.F.
Weber. 1982. Fast-growing rhizobia isolated from root nodules of soybean. Science 215: 1631-1632.
Kingsley, M.T., and B.B. Bohlool. 1983.
Characterization of Rhizobium sp. (Cicer arietinum
L.) by immunofluorescence, immunodiffusion, and intrinsic antibiotic
resistance. Can. J. Microbiol. 29:
518-526
Kondorosi, A., E. Kondorosi, C.E. Pankhurst,
W.J. Broughton, and Z. Banfalvi. 1982.
Mobilization of a Rhizobium meliloti megaplasmid carrying
nodulation and nitrogen fixation genes into other rhizobia and Agrobacterium. Mol. Gen. Genet. 188: 433-439.
Kondorosi, E., Z. Banfalvi, and A.
Kondorosi. 1984. Physical and genetic analysis of a symbiotic
region of Rhizobium meliloti: identification of nodulation genes. Mol. Gen. Genet. 193: 445-452.
Kowalczuk, E., A. Skorupska, and Z.
Lorkiewicz. 1981. Transfer of nodulation ability in Rhizobium
using RP68.45 derived plasmids. Mol.
Gen. Genet. 183: 388-391.
Krol, A.J.M., J.G.J. Hontelez, R.C. Van den
Bos, and A. Van Kammen. 1980.
Expression of large plasmids in the endosymbiotic form of Rhizobium leguminosarum. Nucleic Acid Res. 8: 4337-4347.
Kuykendall, L.D. 1979. Transfer of R
factors to and between genetically marked sublines of Rhizobium japonicum. Appl. Environ. Microbiol. 37: 862-866.
Lamb, J.W., G. Hcmbrecher, and A.W.B.
Johnston. 1982. Plasmid-determined modulation and
nitrogen-fixation abilities in Rhizobium phaseoli. Mol. Gen.
Genet. 186: 449-452.
Leonard, L.T. 1943. A simple assembly
for use in the testing of cultures of rhizobia. J. Bacteriol. 45: 523-527.
Ljunggren, H. and G. Fahraeus. 1961.
The role of polygalacturonase in root hair invasion by nodule
bacteria. J. Gen. Microbiol. 26: 521-528.
Long, S.R., W.J. Buikema, and F.M. Ausubel. 1982.
Cloning of Rhizobium meliloti nodulation genes by direct
complementation of Nod- mutants. Nature 298: 485-488.
Marvel, D.J., G. Kuldau, A.M. Hirsch, J.
Park, J.G. Torrey,and F.M. Ausubel.
1984. Cloning and
characterization of a nodulation locus from Rhizobium parasponium,
p. 691. In C. Veeger and W.E. Newton
(ed.), Advances in nitrogen fixation research, Nijhoff-Junk, The Hague, The
Netherlands.
Masterson, R.V., R.K. Prakash, and A.G.
Atherly. 1985. Conservation of symbiotic nitrogen fixation
gene sequences in Rhizobium japonicum and Bradyrhizobium japonicum. J. Bacteriol. 163: 21-26.
Masterson, R.V., P.R. Russel, and A.G.
Atherly. 1982. Nitrogen fixation ( nif ) genes and large
plasmids of Rhizobium japonicum.
J. Bacteriol. 152: 298-331.
Mathis, J.N., W.M. Barbour, and G.H.
Elkan. 1985. Effect of sym plasmid curing on symbiotic effectiveness in Rhizobium
fredii. Appl. Env. Microbiol.
49: 1385-1388.
Meade, H.M., S.R. Long, G.B. Ruvkun, S.E.
Brown, and F.M. Ausubel. 1982. Physical
and genetic characterization of symbiotic and auxotrophic mutants of Rhizobium
meliloti induced by transposon Tn5 mutagenesis. J. Bacteriol. 149:
114-122.
Morrison, N.A., Y.H. Cen, M.J. Trinick, J.
Shine, and B.G.Rolfe. 1983. Heat curing of a sym plasmid in a
fast-growing cowpea Rhizobium sp. that is able to nodulate legumes and
the nonlegume Parasponia. J.
Bacteriol. 153: 527-531.
Nadler, K.D. 1984.
Transfer of symbiotic and conjugative plasmids to and from fast-growing R.
japonicum, p.712. In. C. Veeger
and W.E. Newton (ed.), Advances 1n nitrogen fixation research. Nijhoff--Junk,
The Hague, The Netherlands.
Noti, J.D., B. Dudas, and
A.A. Szalay. 1985. Isolation and characterization of nodulation
gene from Bradyrhizobium sp. (Vigna) strain IRc78. Proc. Natl. Acad. Sci. 82: 7379-7383.
Nuti, M.P., A.M.
Ledeboer, A.A. Lepidi, and R.A. Schilperoort.
1977. Large plasmids in different Rhizobium species. J. Gen. Microbiol. 100: 241-248.
Nuti, M.P., A.A. Lepidi,
R.K. Prakash, R.A. Schilperoort, and F.C. Cannon. 1979. Evidence for nitrogen fixation (nif) genes
on indigenous Rhizobium plasmids.
Nature 282: 533-535.
Palomares, A., E.
Montoya, and J. Olivares. 1978. Induction of polygalacturonase production in
legume roots as a consequence of extrachromosomal DNA carried by R. meliloti. Microbios 21: 33.
Parijskaya, A.N. 1973.
The effect of acridine orange and mitomycin C on the symbiotic
properties of R. meliloti.
Mikrobiologiya 42: 119-121.
Pilacinski, W.P. and E.L.
Schmidt. 1981. Plasmid transfer within and between
serologically distinct strains of Rhizobium japonicum, using
antibiotic resistance mutants and auxotrophs.
J. Bacteriol. 145: 1025-1030.
Prakash, R.K., P.J.J.
Hooykaas, A.M. Ledeboer, J.W.Kijne, R.A. Schilperoort, M.P. Nuti, A.A. Lepidi,
F. Casse, C. Boucher, J.S. Julliot, and J. Denarie. 1980. Detection,
isolation, and characterization of large plasmids in Rhizobium, p.
139-163. In: W.E. Newton and W.H.
Orme-Johnson (ed.), Proceedings of the III International Symposium on Nitrogen
Fixation, Wisconsin University Park Press, Baltimore.
Prakash, R.K., R.A.
Schilperoort, and M.P. Nuti. 1981. Large plasmids of fast-growing rhizobia:
homology studies and location of structural nitrogen fixation (nif) genes. J. Bacteriol. 145: 1129-1136.
Prakash, R.K., and A.G.
Atherly. 1984. Reiteration of genes involved in symbiotic
nitrogen fixation by fast-growing Rhizobium japonicum. J. Bacteriol. 160. 785-787.
Rolfe, B.G., M.
Djordjevic, K.F. Scott, J.E. Hughes, J. Badenoch Jones, P.M. Gresshoff, Y. Cen,
W.F. Dudman, W. Zurkowski, and J. Shine. 1981.
Analysis of the nodule-forming ability of fast growing Rhizobium
strains, p. 142-145. In: A.H. Gibson
and W.E. Newton (ed.), Current perspectives in nitrogen fixation, Australian
Academy of Sciences, Canberra.
Ronson, C.W. and D.B.
Scott. 1983. Identification, broad host range mobilization and mutagenesis of
a Rhizobium trifolii Sym::R68.45 cointegrate plasmid, p.
177-187. In: A. Puhler (ed.), Molecular
genetics of the bacteria-plant interaction.
Springer, Berlin, Heidelberg, New York.
Rosenberg, C., P.
Boistard, J. Denarie, and F. Casse-Delbart.
1981. Genes controlling early and late functions in symbiosis are
located on a megaplasmid in Rhizobium meliloti. Mol. Gen. Genet. 184: 326-333.
Rosenberg, C., F.
Casse-Delbart, I. Dusha, M. David, and C. Boucher. 1982. Megaplasmids in the plant associated
bacteria Rhizobium meliloti and Pseudomonas solanacearum. J. Bacteriol. 150: 402-406.
Ruvkun, G.B., and F.M.
Ausubel. 1980a. Interspecies homology of nitrogenase
genes. Proc. Nat. Acad. Sci. 77:
191-195.
Sadowsky, M.J. 1983.
Physiological, serological, and plasmid characterization of fast-growing
rhizobia that nodulate soybeans. Ph.D. thesis. University of Hawaii.
Sadowsky, M.J., and B.B.
Bohlool. 1983. Possible involvement of a megaplasmid in
nodulation of soybeans by fast-growing rhizobia from China. Appl. Environ. Microbiol. 46: 906-911.
Sadowsky, M.J., and B.B.
Bohlool. 1985. Differential expression of the pea symbiotic
plasmid pJB5JI in genetically dissimilar backgrounds. Symbiosis 1:125-138.
Schmidt, E.L., R.O.
Bankole, and B.B. Bohlool. 1968. Fluorescent antibody approach to the study
of rhizobia in soil. J. Bacteriol. 95:
1987-1992.
Schofield, P.R., M.A.
Djordjevic, B.G. Rolfe, J. Shine, and J.M. Watson. 1983. A molecular linkage map of nitrogenase and
nodulation genes in R. trifolii.
Mol. Gen. Genet.192: 459-465.
Schofield, P.R., R.W.
Ridge, B.G. Rolfe, J. Shine, and J.M. Watson.
1984. Host-specific nodulation
is encoded on a 14 kb fragment in Rhizcbium trifolii. Plant Mol.Biol. 3: 3-11.
Scholla, M.H., and G.H.
Elkan. 1984. Rhizobium fredii sp. nov., a fast-growing species
that effectively nodulates soybeans.
Int. J. Syst. Bacteriol. 34: 484-486.
Schwinghamer, E.A. 1980.
A method for improved lysis of some Gram-negative bacteria. FEMS Microbiol. Lett. 7: 157-162.
Scott, D.B., C.B. Court,
C.W. Ronson, K.F. Scott, J.M. Watson, P.R. Schofield, and J. Shine. 1984.
Organization of nodulation and nitrogen fixation genes on a Rhizobium
trifolii symbiotic plasmid. Arch. Microbiol. 139: 151-157.
Scott, D.B., and C.W.
Ronson. 1982. Identification and mobilization by cointegrate formation of a
nodulation plasmid in Rhizobium trifolii. J. Bacteriol. 151: 36-43.
Simon, R., U. Priefer,
and A. Puhler. 1983. Vector plasmds for in vivo and in vitro
manipulation of Gram-negative bacteria, p. 98-106. In: Molecular genetics of the bacteria-plant interaction. Springer-Verlag, Berlin, Heidelberg.
Truchet, G., C.
Rosenberg, J. Vasse, J. Julliot, S. Camut,and J. Denarie. 1984.
Transfer of Rhizobium meliloti pSym genes into Agrobacterium
tumefaciens: hostspecific nodulation by a typical infection. J. Bacteriol. 157: 134-142.
Verma, D.P., and A.
Long. 1983. The molecular biology of Rhizobium-legume symbiosis. Int. Rev. Cyt. Supp. 14: 211-43.
Vincent, J.M. 1970.
A manual for the practical study of root-nodule bacteria. IBP Handbook No. 15. Blackwell Scientific Publications, Oxford.
Zurkowski, W. 1981.
Conjugal transfer of the nodulation conferring plasmid pWZ-2 in R.
trifolii. Mol. Gen.Genet. 181:
522-524.
Zurkowski, W. 1982.
Molecular mechanism for loss of nodulation properties of Rhizobium
trifolii. J. Bacteriol. 150:
999-1007.
Zurkowski, W., M. Hoffman,
and s. Lorkiewicz. 1973. Effect of acriflavin and sodium dodecyl
sulphate on infectiveness of Rhizobium trifolii. Acta Microbiol.
Polon. 5: 55-60.
Zurkowski, W., and Z.
Lorkiewicz. 1976. Plasmid deoxyribonucleic acid in Rhizobium
trifolii. J. Bacteriol. 128:
481-484.
Zurkowski, W., and Z.
Lorkiewicz. 1978. Effective method for the isolation of
non-nodulatiog mutants of Rhizobium trifolii. Gen. Res. 32: 311-314.